
Research
Two-Component Signal Transduction Systems (TCSTS).
Myxococcus xanthus is a Gram negative soil-dwelling organism renowned for its co-operative social behaviour. M. xanthus predates other members of the soil microfauna in a social phenomenon reminiscent of wolf-pack hunting. When nutrients and prey species become scarce, a population of many thousand M. xanthus cells aggregate and develop morphologically complicated structures called fruiting bodies. Within these fruits, a subset of cells differentiate into resistant cell forms called myxospores.
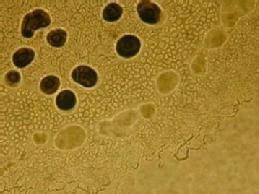
A population of Myxococus xanthus fruiting on an agar surface.
Such behaviours require precise cellular responses to environmental changes, and these environmental changes include interactions with other cells within the population. The full genome sequence of M. xanthus has revealed an unparalleled number of signalling pathways in this organism. A particularly abundant type of signalling pathway in M. xanthus is the two-component system (TCS). In a typical system the first component is a histidine protein (or sensor) kinase while the second component is a response regulator protein. The sensor kinase senses environmental change through an 'input' domain, which triggers autophosphorylation of its 'transmitter' domain. Subsequently the phosphoryl group is transferred to the 'receiver' domain of the partner response regulator, leading to an altered function of its 'output' domain and causing a response to the initial stimulus.
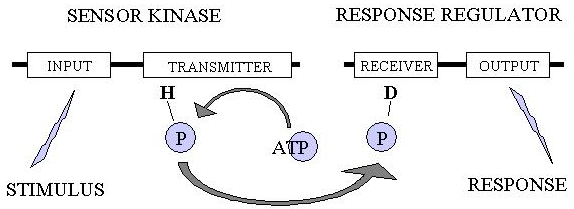
A typical two-component system.
Many of the 255 TCS proteins in the M. xanthus genome are orphaned (isolated from other TCS genes) and there is consequently no clue as to which partner proteins they communicate with. My initial approach is to map the interactions between all transmitter and receiver domains encoded in the M. xanthus genome to define TCS signalling partnerships. As a consequence of the approach, I am also able to measure cross-communication between different systems and to determine whether systems act independently or operate together as networks. Initial results imply a great deal of communication between different TCS pairs, and hints at the existence of complicated signalling networks.
Experimental work is complemented by more theoretical analyses. Statistical models are being developed to enable predictions of TCS partnerships to be made from genomic sequence. As well as identifying potential partners for orphan TCS proteins, such tools also allow signalling pathways to be deduced. The properties of predicted and experimentally determined signalling networks will then be analysed by mathematical modelling. My long term goals are to understand TCS regulation at the system level in the myxobacteria, and to learn how system properties change during evolution.